

Sexuelle Selektion - kritische Beurteilung
- 13. Mai
- 27 Min. Lesezeit
Aktualisiert: 21. Juni
Einleitung:
Sexuelle Selektion ist eine wichtige Annahme der Evolutionsbiologie. Sie wird meist als Erklärung herangezogen, wenn ein Merkmal keinen erkennbaren Überlebens- oder Fruchtbarkeitsvorteil bietet und deshalb nicht durch natürliche Selektion erklärbar ist.
Obwohl sexuelle Selektion ein häufiger Bestandteil evolutionstheoretischer Erklärungen ist, scheint es hierzu von schöpfungswissenschaftlicher Seite vergleichsweise wenig Veröffentlichungen zu geben. – auf Deutsch scheinbar bislang keine einzige. Sie wird zwar oft erwähnt, doch dabei eher als ein Unterthema behandelt. Detaillierte Darstellungen oder Diskussionen mit dem Fokus auf dieses Thema sind selten. Der vorliegende Artikel soll diese Lücke schließen. Er basiert auf wenigen englischsprachigen schöpfungswissenschaftlichen Quellen und eigener Recherche in evolutionstheoretischer populärwissenschaftlicher Literatur und Fachliteratur.
Dieser Artikel ist in zwei unabhängige Teile gegliedert. Der erste und kürzere Teil richtet sich an interessierte Laien, der zweite und längere Teil richtet sich an fortgeschrittene Leser, die mehr Detailtiefe wünschen.
Interessierte Laien:
Es gibt viele Tierarten, bei denen ein Geschlecht um das andere für die Paarung wirbt – meist das Männchen um das Weibchen. Dabei kommen oft Merkmale, bzw. Verhaltensweisen zum Einsatz, die ein Geschlecht vom anderen unterscheiden, aber nicht am Geschlechtsakt beteiligt sind. Sie werden als sekundäre Geschlechtsmerkmale bezeichnet. Durch diese Merkmale / Verhaltensweisen soll ein Partner gewonnen und so der Fortpflanzungserfolg gesteigert werden
Dabei wird zwischen 2 Kategorien unterschieden:
Merkmale / Verhaltensweisen, die nur dem anderen Geschlecht imponieren sollen.
Merkmale / Verhaltensweisen, die dem Wettkampf um Weibchen zwischen Männchen dienen.
Zu Ersterem zählt u.a. der Fregattvogel, der seinen leuchtend roten Kehlsack aufbläst, um Weibchen anzulocken. Der japanische Kugelfisch kreiert zur Balz im Verlauf von 7-9 Tagen ein kreisrundes, symmetrisches Kunstwerk im Sand, das er darüber hinaus mit Muscheln verziert. Es hat einen Durchmesser von bis zu zwei Metern und in der Mitte werden dann die Eier abgelegt und befruchtet.
Zu Letzterem gehört bspw. das Hirschgeweih, mit dem Hirsche den Rivalenkampf um die Paarung mit den Hirschkühen austragen.


Soweit die beobachtbaren und unumstrittenen Tatsachen. Umstritten ist dagegen, wie plausibel die Entstehung und Durchsetzung dieser Merkmale durch sexuelle Selektion erklärt werden kann.
Merkmale, die lediglich der sexuellen Selektion dienen sollen, haben meist keinen direkten Überlebens- oder Fruchtbarkeitsvorteil. Dazu ist ihre Ausbildung und Aufrechterhaltung für den jeweiligen Körper zum Teil mit hohem Energieaufwand verbunden. Diese Energie fehlt dann in anderen, für das Überleben unmittelbar wichtigen Körpersystemen.
Deshalb sollten sich unnötig energieaufwendige oder anderweitig für das Überleben nachteilige Merkmale aufgrund der natürlichen Selektion eigentlich nicht dursetzen. Als einzige Erklärung bleibt Vertretern der Evolutionslehre dann noch, dass solche Merkmale den Fortpflanzungserfolg des jeweiligen Individuums so stark fördern, dass sie dennoch weiter mitvererbt und nicht ausselektiert werden.
Die folgenden Ausführungen beziehen sich nur auf die sexuelle Selektion von Merkmalen, die dem anderen Geschlecht imponieren sollen, da diese Form am bekanntesten ist und gerne als Erklärung für Merkmale gebraucht wird, die aufgrund ihrer (unnötigen) Schönheit auch als Schöpfungsindiz angesehen werden.
Daraus ergibt sich das Hauptproblem, dass ein neues Merkmal nur dann sexuell selektiert werden kann, wenn das andere Geschlecht dieses Merkmal bevorzugt. Doch woher sollen Weibchen eine Präferenz für ein neues Merkmal haben, welches es bis vor kurzem noch nicht gab? Auch die Bevorzugung eines Merkmals müsste sich erst entwickeln, das wäre aber erst nach der Entstehung des zu bevorzugenden Merkmals möglich. Doch wie kann ein nutzloses Merkmal in der Population bestehen, solange es von den Weibchen noch nicht bevorzugt wird? Es würde in der Zwischenzeit viel eher aufgrund von Nutzlosigkeit oder durch die ungehinderte Ansammlung schadhafter Mutationen ausselektiert werden. Die alternative Möglichkeit wäre eine "Vorprogrammierung" der weiblichen Bevorzugung, doch dies verschiebt den Erklärungsbedarf auf die Entstehung der Programmierung, die an sich für die Existenz eines Programmierers spricht.
Evolutionsvertreter könnten hier höchstens noch eine zufällige paralle Entwicklung von Merkmal und Präferenz annehmen, doch abgesehen von der hohen Unwahrscheinlichkeit dieses Szenarios, käme man auch hier nicht um eine Zwischenzeit ohne positiven Selektionsdruck rum, da die genetische Grundlage der Vorliebe ja ebenfalls ihre Zeit zur ausreichenden Entwicklung bräuchte.
Fazit:
Es ist eine beobachtbare Tatsache, dass manche geschlechtsspezifischen Merkmale am Fortpflanzungserfolg ihres Trägers beteiligt sind. Dieser Sachverhalt erklärt aber nicht ihre erstmalige evolutive Entstehung. Vor allem die Tatsache, dass die Bevorzugung eines Merkmals vor der Entstehung des Merkmals keinen Sinn ergibt, ein neues Merkmal ohne anfängliche Bevorzugung aber keinen Vorteil hat und daher nicht erhalten bleibt, ist ein erhebliches Problem für das Konzept der sexuellen Selektion.
Im Schöpfungsmodell kann sexuelle Selektion hingegen als erhaltende Kraft fungieren, um geschaffene schöne, aber in anderer Hinsicht nachteilige Merkmale zu erhalten (roter Kehlsack des Fregattvogels) oder bei manchen aufwändigen Verhaltensweisen einfach Ausdruck der Kreativität des Schöpfers sein (aufwändiger Balzplatz beim Japanischen Kugelfisch). Doch sie kann keinen Schöpfer ersetzen.
Fortgeschrittene:
Was ist sexuelle Selektion?
Die Idee der sexuellen Selektion geht bis auf Charles Darwin zurück. Er konnte sich so manches unnötig schöne Merkmal wie das Rad des Pfau oder das Prachtgefieder der Paradiesvögel nicht mit natürlicher Selektion erklären und vermutete eine "weibliche Zuchtwahl" (Darwin 1871; vgl. Witte 2009) für die Fixierung solch schöner Merkmale.
Seitdem wurde sexuelle Selektion unter Wissenschaftlern ausgiebig diskutiert. Während Darwin die sexuelle Selektion noch klar von der natürlichen Selektion trennte, wollte der Mitbegründer der Evolutionstheorie Alfred Russel Wallace sie als Teil der natürlichen Selektion behandeln. Heute wird sie als Unterkategorie der natürlichen Selektion betrachtet; im vorliegenden Artikel wird der Begriff "natürliche Selektion" (im engeren Sinne) allerdings immer dann verwendet, wenn es sich explizit nicht um sexuelle Selektion handelt.
Einige Zitate zur Veranschaulichung dieser Entwicklung:
Shuker & Kvarnemo 2021: "Da unsere Definition anderen Fitnesskomponenten gegenüber agnostisch bleibt, stellt sich die Frage: Wie verortet sie die sexuelle Selektion im Verhältnis zur natürlichen Selektion? [...] Wie oben erwähnt, wurde von Endler (1986) eine besonders nützliche Konzeptualisierung bereitgestellt. Er schlug vor, dass die natürliche Selektion im weiteren Sinne sämtliche denkbaren Fitnesskomponenten umfassen sollte (gewissermaßen als ein vollständiges Kompendium darwinistischer Selektionsmöglichkeiten), während die natürliche Selektion im engeren Sinne Bereiche wie die Viabilitäts- und Fekunditätsselektion [Überlebens- und Fruchtbarkeitsselektion] abdecken sollte, da diese traditionell eher als ‚natürliche Selektion‘ aufgefasst wurden und den Hauptfokus von Darwins Die Entstehung der Arten bildeten." (Hervorhebung hinzugefügt, eigene Übersetzung)
Shuker & Kvarnemo verbinden mit ihrem aktuellen Artikel zur Definition (siehe unten) sexueller Selektion beide Positionen. In der natürlichen Selektion geht es um Fitness. Komponenten der Fitness sind Überlebensfähigkeit, Fruchtbarkeit und der sich gewöhnlich daraus ergebende Fortpflanzungserfolg. Je länger ein Individuum lebt und je fruchtbarer es ist, desto mehr Nachkommen kann es zeugen. Längerfristig ist Fitness aber nur am Fortpflanzungserfolg messbar.
Die von Shuker & Kvarnemo erwähnte traditionelle Auffassung natürlicher Selektion beinhaltet eben diese Gleichung: Überlebensfähigkeit + Fruchtbarkeit => Fortpflanzungserfolg.
Sexuelle Selektion bringt nun einen Widerspruch in diese Rechnung. Sie wirkt sich direkt auf den Fortpflanzungserfolg aus und erhöht diesen, während sie zugleich häufig die Fitnesskomponente der Überlebensfähigkeit reduziert. Das einzelne männliche Individuum ist während seines Lebens gemessen an seiner Überlebensfähigkeit weniger fitt, doch aufgrund seines desto höheren Fortpflanzungserfolges (in potenziell kürzerer Zeit) ist sein Genotyp über Generationen gemessen fitter. Daraus resultierte die Uneinigkeit, ob sexuelle Selektion eigenständig oder ein Teil natürlicher Selektion ist.
Die vorgeschlagene Differenzierung zwischen natürlicher Selektion im engeren und weiteren Sinn löst den Widerspruch. Die engere natürliche Selektion, welche sich direkt auf das Leben eines Individuums auswirkt, steht im Gegensatz zur sexuellen Selektion. Gleichzeitig wirkt sich die sexuelle Selektion durch ihre Beeinflussung des Fortpflanzungserfolges auf die längerfristig messbare weitere natürliche Selektion aus und ist deshalb ein Teil von ihr.
Was dagegen seit Darwin beibehalten wurde, ist die Unterscheidung zweier Mechanismen der sexuellen Selektion: Die intrasexuelle Selektion und die intersexuelle Selektion (Darwin 1871, S. 398; Shuker & Kvarnemo 2021).
Intrasexuelle Selektion bezieht sich meist auf den Wettkampf zwischen Männchen um Weibchen, seltener konkurrieren in manchen Arten auch die Weibchen um die Männchen. Sie soll sekundäre Geschlechtsmerkmale fördern, "die die Konkurrenzfähigkeit eines Geschlechts um den Zugang zum anderen Geschlecht wahrscheinlicher machen", wie es Witte (2009) formuliert.
Zu solchen sekundären Geschlechtsmerkmalen zählt bspw. das Geweih des Hirsches. Es ist energetisch aufwendig zu produzieren und zu erhalten, wobei es für die Verteidigung gegen Raubtiere nicht besonders hilfreich ist, zumal es jährlich abgeworfen wird und neu nachwächst. Zur Verteidigung wären dauerhafte Hörner besser. Das Geweih kann beim Laufen durch den Wald und der Flucht sogar hinderlich sein und es erhöht den Nahrungsbedarf des Hirsches (Bolten 2020). Es dient einzig dem Wettkampf zwischen den Hirschen einer Herde in der Paarungszeit. Der Sieger darf sich mit den Hirschkühen fortpflanzen.
Ein weiteres Beispiel ist das Brustschild der männlichen See-Elefanten. See-Elefanten-Bullen kämpfen blutig um die Vorherrschaft und das alleinige Paarungsrecht am Strand. Sie prallen dabei mit Brust und Hals aufeinander und setzten ihre Eckzähne ein. Eine verdickte Hornhautschicht in diesem Bereich schützt sie vor tödlichen Verletzungen.
Neben sekundären Geschlechtsmerkmalen, die dem Angriff oder der Verteidigung gegen gleichgeschlechtliche Artgenossen dienen, gibt es auch Merkmale, die der Einschüchterung oder Abschreckung dienen, sodass sich ein anderes Männchen ohne Kampf zurückzieht.
Hier kann wieder der Hirsch mit seinem Röhren in der Brunftzeit genannt werden. Die Tonhöhe und die Ausdauer dieser Rufe erlaubt einem Artgenossen die Einschätzung der körperlichen Stärke, so dass dieser entscheiden kann, wann ein Kampf lieber zu vermeiden ist.
Intersexuelle Selektion beschreibt das Bemühen eines Individuums eines Geschlechtes ein Individuum des anderen Geschlechtes zu beeindrucken, sodass dieses es für die Paarung auswählt. Meist wirbt hier das Männchen um das Weibchen (Clutton-Brock & Parker 1992, S. 438; vgl. Bergman 1996, S. 217). Das kann sowohl mit Hilfe optischer Merkmale geschehen, als auch durch bestimmte Verhaltensweisen.

Der Lockenvogel-Paradiesvogel (Lophorina niedda) vereint beides. Das Männchen verfügt sowohl über einen schimmernd blauen Federkranz, auch die Fähigkeit einen Balztanz aufzuführen. Zur Balz bildet er mit seinen schwarzen Federn eine Sichelform, wobei etwa auf Brusthöhe in einer waagerechten Linie sein Brustschild aus blauen Federn und darüber seine blauen "Augen" aus blauen Kronenfedern hervorstechen. In dieser Haltung tänzelt er um das Weibchen herum (Scholes & Laman 2018).

Ein sehr bekanntes und von Evolutionsvertretern gern genutztes Beispiel sind die Schwanzfedern des männlichen Pfaus. Während die Pfauenhenne unauffällig braun gefärbt ist, hat der männliche Pfau ein wunderschönes, bunt schillerndes Gefieder. Im "Kampf ums Überleben" hat das eigentlich nur Nachteile. Die langen Schwanzfedern sind energetisch aufwändig zu produzieren und zu erhalten, ohne dabei funktionell sinnvoll zu sein und für Fressfeinde ist das Männchen viel leichter zu entdecken und zu fangen. Die Pfauen schlagen ihr Rad hauptsächlich zur Balz auf.
Als einzige Erklärungsmöglichkeit im Rahmen der Evolutionstheorie bleibt die Präferenz der Pfauenhenne für diese Schönheit.
Sexuelle Selektion umfasst also nicht nur das allgemein bekannte Balzverhalten, durch welches das andere Geschlecht gewonnen werden soll, sondern auch den Wettkampf um das andere Geschlecht. Letzterer ist vor allem bei Arten mit polygynem Paarungssystem anzutreffen, in dem wenige Männchen sich mit vielen Weibchen paaren und so der Fortpflanzungserfolg der übrigen Männchen gemindert wird (Witte 2009).
Die intrasexuelle Selektion endet allerdings nach heutigem Verständnis nicht mit der Paarung. Wenn ein Weibchen sich mit mehreren Männchen gepaart hat, kann unter Umständen (nach Wigby & Chapman 2004) zwischen den verschiedenen Spermien noch ein "Wettlauf", bzw. Wettbewerb zu der Eizelle stattfinden – die sogenannte Spermienkonkurrenz.
Dabei kann es u. a. auf die produzierte Spermienmenge (mehr Spermien – höhere Wahrscheinlichkeit auf Befruchtung) und die Schnelligkeit der Spermien ankommen (Simmons & Wedell 2020). Manche Arten produzieren auch nicht-befruchtende Spermien, die den Spermien eines anderen Männchens den Zugang zur Eizelle blockieren (Wigby & Chapman 2004). Spermienkonkurrenz ist ein recht großes Forschungsfeld. Ein Übersichtartikel von Simmons & Wedel (2020). fasst die Forschungsergebnisse von 50 Jahren zusammen.
Vor dem genannten Hintergrund formulieren Shuker & Kvarnemo (2021) ihre aktuelle Definition der sexuellen Selektion recht umfassend:
"Sexuelle Selektion ist jede Form der Selektion, die aus Fitnessunterschieden hervorgeht, welche mit nicht-zufälligem Erfolg im Wettbewerb um den Zugang zu Gameten für die Befruchtung verbunden sind."
Diese Definition umfasst sowohl intersexuelle als auch intrasexuelle Merkmale sowie durch die Formulierung "Gameten" (Geschlechtszellen) auch die Spermienkonkurrenz.
Kritische Beurteilung:
1) Merkmal oder Präferenz zuerst?
In der schöpfungsorientierten Literatur wird bei intersexueller Selektion meist kritisch angemerkt, dass unklar ist, sie sich die Präferenz der Weibchen herausgebildet hat. In evolutionstheoretischen Beschreibungen wird diese oft unkommentiert als gegeben vorausgesetzt, doch ihre Herkunft erfordert eine Erklärung. Eine bereits vorhandene, also „vorprogrammierte“ Präferenz würde einen vorausplanenden Programmierer erfordern, denn Evolutionsmechanismen können bekanntermaßen nicht programmieren.
Wenn aber die Gene, die bei der Präferenz eines Merkmals eine Rolle spielen, erst nach dem Auftreten des bevorzugten Merkmals entstehen (was sicher nicht mit einer Mutation getan wäre), fehlt für dieses Merkmal in der Zwischenzeit die stabilisierende Selektion, die seinen Erhalt in der Population ermöglichen würde. Man könnte an der Stelle zwar einwenden, dass überflüssige Merkmale ohne Nachteil erhalten bleiben können, doch muss man hier zwei Gegenargumente beachten:
1. Nicht wenige sekundäre Geschlechtsmerkmale wie das Schwanzgefieder der Pfauen sind durchaus von Nachteil, weil sie energetisch aufwändig sind bzw. die Gefahr erhöhen, dass das Tier von Fressfeinden entdeckt wird.
2. Darüber hinaus sind über 99 % der Mutationen schädlich (Natterer 2010). Ein nicht durch Selektion geschütztes Merkmal würde daher ungehindert schädliche Mutationen ansammeln. Bergman (2005) führte dazu eine eindrückliche Stichprobe durch. Er durchsuchte 2005 alle in den Jahren von 1966 bis 2003 publizierten Studien mittels Computersuche nach positiven Mutationen. Von 16 Millionen beschriebenen Mutationen waren nur 186 vorteilhaft. Das sind 0,0011625 %.
Typischerweise ist also zu erwarten, dass erstens ein neues funktionsloses sekundäres Geschlechtsmerkmal ohne sofortige Bevorzugung durch Weibchen entweder aufgrund seiner Nachteile ausselektiert würde, bevor es überhaupt nennenswert ausgebildet ist. Zweitens ist erwartbar, dass sich ungehindert schädliche Mutationen ansammeln und das in Entstehung begriffene Merkmal wieder aus der Population verschwindet. Dasselbe gilt, wenn man annimmt, dass Gene, die für die Präferenz der Weibchen erforderlich sind, zeitlich vor dem Auftreten des bevorzugten Merkmals entstanden sein könnten (vgl. Burgess 2023).
Um dieses Problem zu umgehen, entwickelte Ryan (1990) die Sensory-exploitation-Hypothese (vgl. Witte 2009). Sie besagt, dass Weibchen eine latente Präferenz besitzen können, bevor beim Männchen ein entsprechendes sexuelles Merkmal entstanden ist. Dies wird mit weiteren Selektionsfaktoren erklärt. Wenn z.B. ein Weibchen eine bestimmte Frucht besonders gerne frisst und ein Männchen ein Merkmal mit der gleichen Farbe wie die der Frucht entwickelt, soll sich die Bevorzugung auf das Merkmal übertragen können.
Tatsächlich kann in Einzelfällen eine Übereinstimmung von weiblicher Präferenz für bestimmte Merkmale der Umgebung und männlichen Balzmerkmalen empirisch nachweisbar sein.

Weibliche Laubenvögel bevorzugen bei der Futterwahl bestimmte Farben, wie in einem Zoo experimentell nachgewiesen wurde. Die Männchen bauen zur Balz beeindruckende Lauben und verzieren sie u. a. mit Beeren, Blüten oder Gegenständen menschlichen Ursprungs. In besagtem Experiment stimmte die Farbpräferenz der Weibchen für Futter mit der Wahl der Farben bei den Männchen für die Verzierung der Laube mittels Trauben überein (Madden & Tanner 2003; vgl. Witte 2009).
Ein weiteres Beispiel sind Guppys. Die Weibchen dieser Fischart bevorzugen Männchen mit großen orangen Flecken. Gleichzeitig bevorzugen beide Geschlechter orangefarbene Nahrung (Rodd et al. 2002).
Diese Hypothese mag zwar in Einzelfällen einen Erklärungsansatz bieten, taugt aber nicht als allgemeingültige Erklärung, da bei vielen intersexuellen Merkmalen diese Übereinstimmung nicht nachweisbar ist. Die prächtigen Schwanzfedern des männlichen Pfaus weisen beispielsweise keinerlei Ähnlichkeiten zur Nahrung (Samen, Früchte, Blüten, Insekten, kleine Wirbeltiere) oder sonstigen Präferenzen der Pfauen auf.
Darüber hinaus ist eine Übereinstimmung von anderweitiger und sexueller Präferenz auch schöpfungstheoretisch deutbar. So könnte ein Schöpfer auch von Beginn an ein schönes sekundäres Geschlechtsmerkmal auf eine anderweitige Präferenz abstimmen, um so die Erhaltung eines in anderer Hinsicht nachteiligen Merkmals zu gewährleisten.
2) Fixierung trotz Selektionsnachteil?
Sekundäre Geschlechtsmerkmale, die einerseits energieaufwändig sind, ohne die Körperfunktionen zu verbessern, andererseits das Individuum für Fressfeinde sichtbarer machen, die Flucht erschweren oder den Nahrungsbedarf erhöhen, sind für das Überleben von Nachteil. Es wäre also zu erwarten, dass Männchen mit solchen Merkmalen tendenziell weniger Nachkommen erzeugen können als Männchen ohne diese Merkmale.
Warum überwiegt die natürliche Selektion im engeren Sinn also nicht die sexuelle Selektion und verhindert nicht die Entstehung sekundärer Geschlechtsmerkmale? Und warum bevorzugen Weibchen überhaupt derart benachteiligte Männchen?
Zur Beantwortung der ersten Frage wurde von Fisher (1930) ein sogenannter "Runaway-Prozess" vorgeschlagen. Schmuckmerkmal und Präferenz widersetzen sich demnach durch Selbstverstärkung der natürlichen Selektion im engeren Sinn. Da Weibchen ein bestimmtes Merkmal bevorzugen und sich nur mit Männchen mit diesem Merkmal fortpflanzen, sind die Gene für Merkmal und Präferenz gekoppelt.
Wenn sich bei Entstehung der weiblichen Bevorzugung das Schmuckmerkmal bereits (entgegen jeder Wahrscheinlichkeit, siehe Kritikpunkt 1) in einer ausreichenden Anzahl von Männchen verbreitet hat, können sich Individuen mit diesem Merkmal und mit dieser Präferenz ohne Inzuchtprobleme im Vergleich zur übrigen nicht-wählerischen Population überproportional vermehren, sodass nicht-wählerische Weibchen und nicht bevorzugte männliche Merkmale nach und nach verdrängt werden. Es wird postuliert, dass sich Merkmalsausprägung und Bevorzugung immer mehr "aufschaukeln" (vgl. Witte 2009). Gestoppt wird dieser Prozess erst von der natürlichen Selektion, wenn die Nachteile zu gravierend werden.
Die Runaway-Hypothese setzt erst nach Entstehung von Merkmal und Präferenz an und ist deshalb in dieser Phase plausibel. Die Lösung der eigentlichen Probleme (Warum wird ein nachteiliges Merkmal überhaupt bevorzugt? Entstand das Merkmal oder die Präferenz zuerst?) wird vorausgesetzt, wenn der Runaway-Prozess beginnt. Denn dieser bewirkt nur eine Verstärkung. Dieser Verstärkungsprozess würde nur dann problematisch, wenn die Ausgangspopulation so klein wäre, dass es zu Inzuchtproblemen kommt. Das ist aus Perspektive der Evolutionstheorie aber unwahrscheinlich, wenn man postuliert, dass das Merkmal bereits einige Zeit vor Entstehung der Präferenz in der Population verbleiben konnte.
Die Runaway-Hypothese beansprucht somit gar nicht, die eigentlichen Probleme der Hypothese der sexuellen Selektion zu lösen. Darüber hinaus ist sie auch auf das Schöpfungsmodell anwendbar: Nämlich als erhaltender Mechanismus geschaffener Extravaganz gegenüber nach dem Sündenfall auftretenden weniger nachteilhaften Vereinfachungen.
Zum Lösen des zweiten Problems, nämlich warum Weibchen überhaupt benachteiligte Männchen bevorzugen, gibt es sogenannte "Gute-Gene-Hypothesen". Das ist ein Sammelbegriff für mehrere Hypothesen. Eine der bekanntesten ist die von Zahavi (1975) entwickelte Handicap-Hypothese (vgl. Witte 2009). Er deutete die nachteiligen Schmuckmerkmale der Männchen als ehrliche "Qualitätsanzeiger". Ein Männchen, das trotz eines nachteiligen Merkmals überleben kann, hat offensichtlich gute andere Gene, bzw. ist entsprechend fit. Die Weibchen würden daher solche Männchen bevorzugen.
So populär die Handicap-Hypothese heute auch ist, sie hat und hatte auch Vertreter der Evolutionslehre als Kritiker. Kirkpatrick (1986) kritisierte, dass wenn die Hypothese zuträfe, es zu einer ernsthaften Verringerung der Überlebensrate in der Population führen würde und die weibliche Partnerpräferenz daher eine andere selektive Kraft beinhalten müsste.
Grose (2011) argumentierte, dass die Akzeptanz der Handicap-Hypothese durch theoretische Modelle gefördert würde, nicht jedoch durch empirische Befunde (vgl. Woetzel 2017).
Nach Woetzel (2017) ist letzteres vor allem deshalb herausfordernd, weil einige empirische Studien "die vom Handicap-Mechanismus postulierten signifikanten Signalkosten nicht nachweisen konnten [Kane & Zollman 2015]. Bei einer Reihe von „Vorzeigearten" [„poster-child species“] konnten die Vorhersagen der Theorie der sexuellen Selektion nach rigorosen Tests nicht bestätigt werden" (Roughgarden & Akçay 2010; eigene Übersetzung, Hervorhebung hinzugefügt).
Woetzel (2017) kritisiert ebenfalls die postulierte Kausalität zwischen nachteiligem Schmuckmerkmal und „guten“ Genen in der Handicap-Hypothese. Könnte man ein nachteiliges Merkmal nicht auch mit entsprechend „schlechten“ Genen in Verbindung bringen? Bei Merkmalen ist es zudem oft nicht so dramatisch, dass eine nachteilige Variante sofort ausselektiert würde, es geht normalerweise viel mehr um leichte Vorteile. Daher bleiben laut Woetzel (2017) leicht nachteilige Merkmale in der Population erhalten, auch ohne ausgleichende „gute“ Gene. In diesem Fall fielen dann aber „gute“ Gene als Grund der weiblichen Bevorzugung weg.
Doch nehmen wir dennoch einmal an, es bestünde tatsächlich eine Kausalität zwischen nachteiligem Schmuckmerkmal und "guten" Genen und ein Weibchen würde ein solches neues Merkmal bevorzugen (bspw. längere Schwanzfedern mit den „Augen“ beim Pfau). Das Merkmal und seine Bevorzugung durch die Weibchen breiten sich dann im Laufe der Generationen unter den Individuen der lokalen Population aus und mit ihnen auch der (Fitness-)Nachteil – im Fall des Pfaus wäre das die bessere Sichtbarkeit für Fressfeinde, der erhöhte Energiebedarf und die Beeinträchtigung bei der Fortbewegung. Darüber hinaus ginge das auf Kosten anderer wichtiger Merkmale, wie etwa Tarnung oder größerer Körpergröße (letzteres mangels ausreichender Energieverfügbarkeit).
Immer größere und schönere Schwanzfedern müssten über viele Generationen selektiert worden sein, "bevor das hochgradig nuancierte System der ehrlichen Signalgebung etabliert und vorteilhaft wurde" (Woetzel 2017). Solange die Federn und ihr Muster noch keine Signalwirkung entwickelt hatten, hätte es keinen Selektionsdruck gegeben. Bleiben wir beim Beispiel einer Pfauenpopulation: Eine Pfauenpopulation, deren Männchen immer mehr ein derart nachteiliges Merkmal entwickeln, wäre im Vergleich zu benachbarten Populationen deutlich im Nachteil im allgemeinen Kampf ums Überleben.
Da benachbarte Populationen selten isoliert bleiben, würden sich die in der natürlichen Selektion erfolgreicheren Populationen schneller ausbreiten, sich mit der benachteiligten Pfauenpopulation vermischen und so letztlich zum Verschwinden des nachteiligen Merkmals der längeren Schwanzfedern führen.
Számadó et al. (2026) gehen besonders hart mit der Handicap-Hypothese ins Gericht. „Das HP [Handicap-Prinzip] ist tot“, lautet ihr Urteil in einer Zwischenüberschrift. Sie führen theoretische Modelle an, die das Handicap-Prinzip widerlegen, indem sie zeigen, „dass die postulierten Handicap-Kosten sowohl unnötig als auch unzureichend sind, um Ehrlichkeit [d. h. Merkmale, die tatsächlich Fitness widerspiegeln] zu erklären.“
Darüber hinaus seien die theoretischen Modelle zur Unterstützung der Handicap-Hypothese falsch interpretiert worden. Sie stellen ferner in Übereinstimmung mit vorherigen Kritikern fest, dass es trotz Jahrzehnten der empirischen Forschung keine eindeutigen Belege für die Handicap-Hypothese gibt, während es zugleich viele widersprechende Befunde gibt. Sie betonen, dass das Feststellen einer Korrelation von der Ehrlichkeit und der Kostspieligkeit eines Merkmals nicht gleichbedeutend mit einer Kausalität zwischen diesen ist.
Für eine ausführliche kritische Auseinandersetzung mit der Handicap-Hypothese siehe Woetzel (2017) und Számadó et al. (2026).
Nicht zuletzt ist eine viel grundlegendere Kritik angebracht. Zahavi (1975) schreibt:
"Ich gehe davon aus, dass sexuelle Selektion wirksam ist, weil sie die Fähigkeit des selektierenden Geschlechts verbessert, die Qualität des selektierten Geschlechts zu erkennen." (Hervorhebung hinzugefügt, eigene Übersetzung)
Diese von der Handicap-Hypothese postulierte Fähigkeit setzt zwei Grundannahmen voraus:
Die Weibchen sind kognitiv in der Lage einen Zusammenhang von neuem Merkmal und Nachteil zu erkennen und sie können daraus auf eine gute Fitness des Männchens schließen.
Den Weibchen ist ein möglichst fitter Paarungspartner überhaupt wichtig.
Nur wenn diese beiden Voraussetzungen gegeben sind, hat die Handicap-Hypothese überhaupt eine Chance sich zu bewähren. Vertreter dieser Hypothese müssten also zunächst den Nachweis bringen, dass Tiere kognitiv dazu fähig sind und waren, den abstrakten Zusammenhang zwischen neuem Merkmal, daraus resultierendem Nachteil und der anschließenden Folgerung auf gute Fitness zu erkennen und dass ihnen diese Fitness ihres Partners überhaupt wichtig ist.
1) Ohne vorherige Erfahrungen ein neues Merkmal an einem Artgenossen als Ursache für einen Überlebensnachteil zu erkennen und daraus auf eine gute Fitness schließen zu können, stellt ein abstraktes und neutrales Kausalverständnis (die Wirkung betrifft einen nicht selber) dar. Derartige Gedankengänge sind für Menschen zwar kein Problem, doch Vertreter der Handicap-Hypothese versäumen hier, dies auch für die (deutlich) weniger intelligenten Tiere nachzuweisen.
Nur vergleichsweise wenige Arten sind dazu fähig, Kausalzusammenhänge zu verstehen, noch weniger, wenn nicht keine Art, können neutrale Kausalzusammenhänge erkennen, also Zusammenhänge, die nicht direkt sie selbst betreffen (Schloegl & Fischer 2017). Bedenkt man außerdem, dass hypothetische evolutive Vorstufen auch kognitiv weniger weit entwickelt gewesen sein müssten, scheitert die Handicap-Hypothese in den meisten Fällen schon an den kognitiven Fähigkeiten der Weibchen.
2) Bei vielen Arten sind die Männchen nicht an der Aufzucht des Nachwuchses beteiligt. Warum sollte das Weibchen also Wert auf ein fittes Männchen legen? Es folgt ja letztlich nur seinem Sexualtrieb und ist nach der Paarung nicht mehr auf dieses angewiesen. Argumentiert man, dass ein fitterer Partner auch fittere Nachkommen bedeutet, setzt man wieder kognitive Kausal-Verständnis-Fähigkeiten voraus, die erstmal nachzuweisen wären. Denn der längerfristige „objektive“ Selektionsvorteil eines fitteren Vaters erklärt nicht die Wahlentscheidung des Weibchens.
Letztlich werden den Weibchen logisch-kausale Gedankengänge unterstellt, die für den intelligenten Menschen zwar selbstverständlich sind – nicht jedoch bei Tieren.
3) Sexuelle Nötigung:
Von Männchen erzwungene oder genötigte Kopulation ist im Tierreich keine Seltenheit, wurde aber häufig nicht genauer in evolutionäre Hypothesen miteinbezogen. So schreiben bspw. Head & Brooks:
Die Bedeutung der Männchen-Männchen-Konkurrenz, unerwünschter Kopulationen und sexueller Belästigung wurde jedoch ignoriert. (Head & Brooks 2006, S. 517; Hervorhebung hinzugefügt.)
Tatsächlich ist sie recht verbreitet (Smuts & Smuts 1993) und bei Säugetieren in immerhin fünf Ordnungen (Primaten, Paarhufer, Raubtiere, Unpaarhufer und Wale) nachgewiesen (Cassini 2020), darüber hinaus auch bei Vögeln (z.B. Brennan et al. 2007), Fischen (z.B. Head & Brooks 2006) und Reptilien (z.B. Lee & Hays 2004).
Bei erzwungener Paarung spielen sekundäre Geschlechtsmerkmale des Männchens keine Rolle mehr und somit kann sie die Wirkung der weiblichen sexuellen Selektion schwächen oder sie zumindest verkomplizieren (Bergman 2004).
Paradebeispiel Pfau:
Der männliche Pfau dient als Paradebeispiel für sowohl unnötige Schönheit, als auch sexuelle Selektion. Das kann auf den ersten Blick für Verwirrung sorgen. Zeugt der Pfau mit seiner enormen Schönheit nun von einem Schöpfer mit Sinn für Schönheit oder ist er das Paradebeispiel weiblicher Partnerwahl?
Zunächst zu den offensichtlichen Daten: Das Rad des männlichen Pfaus besteht aus etwa 200 einzelnen Schwanzfedern, von denen 170 einen dekorativen Augenfleck bilden; die übrigen 30 Federn werden von einem T-ähnlichen Federbusch gekrönt (Burgess 2023).
Ausgebreitet zu einem Fächer mit einem Winkel von über 180 Grad weisen jeder Augenfleck und jedes „T“ einen gleichmäßigen Abstand zueinander auf. Die kürzeren Federn sind vor den längeren angeordnet, sodass jedes „Auge“ sichtbar ist. Die Federn sind symmetrisch und geometrisch präzise angeordnet, die beiden nachfolgenden Bilder sollen dies veranschaulichen:


Wie sollen ungerichtete und ziellose Evolutionsprozesse eine derart geordnete Geometrie und Musterung hervorbringen? Und wie sollte dies durch sexuelle Selektion möglich sein?
Burgess (2023) hat sich in einem ausführlichen Artikel mit den Schwanzfedern des männlichen Pfau und ihrer Erklärbarkeit durch sexuelle Selektion auseinandergesetzt. Er legt detailliert ihren Aufbau dar:
Die Augenfedern:

Im Gegensatz zu einer normalen Flugfeder verläuft der Federschaft bei einer Augenfeder nicht bis zum Ende der Feder. Er endet etwa in der Mitte, wodurch eine Teilung des Augenmusters in zwei Hälften verhindert wird.
Darüber hinaus ist er im Bereich des Musters dünn, wodurch er weniger auffällt und nicht vom eigentlichen Augenmuster ablenkt, und er weist nahe des Musters eine braune Färbung auf. Letzteres ist ebenfalls für die Schönheit der Feder relevant, da ein weißer Schaft im „Auge“ zu stark auffallen würde.
Von dem Schaft zweigen die Federäste ab, die unterhalb des Musters in größeren Abständen zueinander angeordnet sind. So entsteht ein Kontrast zur Präzision der oberen Federäste, die das Augenmuster bilden. Von den Ästen zweigen wiederum die Federstrahlen ab. Am auffälligsten sind die komplexe Musterung und die leuchtenden Farben.
Die Farben leuchten so schillernd aufgrund der Dünnschichtinterferenz. Dies ist ein optischer Effekt, bei dem Licht in den Federstrahlen je nach Winkel unterschiedlich gebrochen wird und so verschiedene Farben erscheinen.
„Im Augenmuster erscheinen die Strahlen bronzefarben, blau, dunkelviolett und grün. Abseits der Augenregion sind die Strahlen einheitlich grün“ (Burgess 2023). Die Farben sind nur auf der Vorderseite zu sehen, wo sich die Federstrahlen befinden. Die Rückseite ist durch die Äste braun pigmentiert.
Zum Verständnis der Funktionsweise der Dünnschichtinterferenz ist zunächst Kenntnis über die detaillierte Struktur der Schwanzfeder wichtig. Wie auch gewöhnliche Flugfedern verfügen die Schmuckfedern des Pfaus über einen Federschaft, abzweigende Äste und davon wiederum abzweigende Federstrahlen. Die Besonderheit liegt in der Strahlenstruktur:
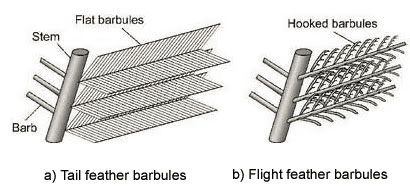
Während die Strahlen bei normalen Federn schmal sind und die Äste nicht überdecken, ähneln sie beim Pfau flachen Bändern, die einander überlappen und so eine eigene Oberfläche auf der Oberseite der Federäste bilden.
Für Dünnschichtinterferenz werden nun eine oder mehrere sehr dünne und transparente Schichten benötigt, die auf einem dunklen Hintergrund liegen. Das transparente Material muss eine zur Wellenlänge des sichtbaren Lichts ähnliche Schichtdicke aufweisen. „Sichtbare Farben haben Wellenlängen zwischen 0,4 und 0,8 µm, und Dünnfilme weisen typischerweise eine Dicke zwischen 0,3 und 1,5 µm auf“ (Burgess 2023).
Darüber hinaus muss der Dünnfilm einen anderen Brechungsindex als den der Luft aufweisen, sodass das Licht beim Durchdringen des Dünnfilms verlangsamt wird. Ein bekanntes Beispiel von Dünnschichtinterferenz ist, wenn eine Öloberfläche auf nassem Untergrund blau und grün schimmert.
Beim Pfau spielt sich die Dünnschichtinterferenz in den drei die Strahlen bedeckenden Keratinschichten ab. Jeder Federstrahl hat eine Breite von etwa 60 µm und eine Dicke von 5 µm. Die Federstrahlen bestehen aus einem Schaumkern von 2 µm Dicke, der auf beiden Seiten von drei Keratinschichten umgeben ist. Die Keratinschichten sind mit einer Dicke von 0,4–0,5 µm extrem dünn.

In den Federstrahlen passiert nun Folgendes: Weißes Licht fällt auf die Keratinschichten und wird sowohl an der Vorderseite als auch der Rückseite des Dünnfilms reflektiert. Das die Keratinschicht(en) durchdringende Licht wird dabei verzögert, sodass auf seinem Rückweg einige der Farbkomponenten des weißen Lichts gegenüber den Lichtwellen, die an der Vorderseite reflektiert wurden, phasenverschoben sind.
Die Phasenverschiebung von zwei Wellenzügen derselben Farbe hat die Aufhebung der Farbe durch destruktive Interferenz zur Folge. Bei weißem Licht ist das Resultat eine Reflexionsfarbe aufgrund der übrigen Farbkomponenten des weißen Lichts.
Das einzige Pigment im Pfauenschwanz ist das Melanin in den Federästen. Durch ihre braune Farbe bieten sie einen guten dunklen Hintergrund für die Dünnschichtinterferenz. Die unterschiedlichen Farben im Augenmuster sind das Ergebnis winziger Änderungen in der Dicke der Keratinschichten, welche dafür auf etwa 0,05 µm genau sein muss.
Die Federstrahlen sind damit optimal konstruiert:
Eine optimale Dicke der Keratinschichten für die Erzeugung der leuchtenden Dünnschichtfarben.
Die dunkelbraune Hintergrundfärbung verhindert, dass Licht durch die Feder strahlt.
Die drei Keratinschichten fügen mehrere Lichtkomponenten hinzu und tragen so zur Brillanz der Farben bei.
Die Strahlen sind in Längsrichtung leicht gekrümmt. Dadurch werden leicht unterschiedliche Farben vermischt, was eine Weichzeichnung der in den Keratinschichten sichtbaren Farben zur Folge hat.
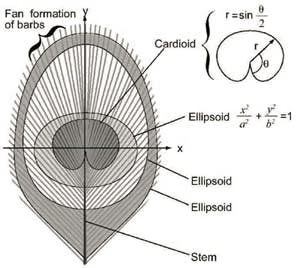
Das Augenmuster ist aufgrund seiner abgerundeten Formen besonders schön. „Die ‚Pupille‘ des Auges wird durch eine dunkelviolette Kardioide [Herzlinie] gebildet und die ‚Iris‘ durch ein blaues Ellipsoid. Diese Formen befinden sich innerhalb eines spitz zulaufenden bronzefarbenen Ellipsoids, das von einem oder zwei grünen Rändern umgeben ist“ (Burgess 2023).
Es handelt sich um ein „digitales Muster“, da es sich aus der kombinierten Wirkung Tausender einzelner Strahlen zusammensetzt. Die voneinander unabhängigen Äste müssen in ihren Positionen exakt angeordnet sein. Auch die Strahlen benachbarter Äste sind für das Augenmuster perfekt aufeinander abgestimmt. Dazu muss die Abfolge der „Farben“ bzw. interferenzwirksamen Strukturen auf jedem Ast im Erbgut vorgeschrieben sein. In der DNA müssen also Zeit- oder Positionsanweisungen vorhanden sein, die für die richtige Keratindicke am richtigen Strahl auf dem richtigen Ast codieren.

Winzige abrupte Änderungen in der Dicke der Keratinschichten der Strahlen entlang eines Astes sorgen für die klaren Grenzen zwischen den Farben. Das ist wichtig, denn graduelle Dickenänderungen würden auch fließende Farbübergänge bedeuten, wodurch das schöne Muster verschwimmen würde. Erstaunlich ist nach Burgess (2023) besonders, dass die Keratindicke entlang eines Astes nicht kontinuierlich zu- oder abnimmt, sondern in einer Richtung beide Änderungen aufweist.
Die T-Federn:

Die langen T-Federn runden das Bild des aufgeschlagenen Pfauenrades perfekt ab. Sie sind zur Augenfeder invers geformt, wodurch das Innenprofil der T-Feder dem Umriss des Augenmusters folgt.
Die T-Federn bilden oftmals einen geschwungenen S-Bogen – eine sogenannte „Ogge-Kurve“ –, was ebenfalls zur Schönheit beiträgt, da sie sowohl konkav als auch konvex ist. Deshalb wird diese Form auch in der Architektur genutzt, beispielsweise beim Kielbogen. Auch diese Form ist bemerkenswerterweise wieder aus vielen einzelnen Federästen zusammengesetzt.
Auch diese Form ist bemerkenswerter Weise wieder aus vielen einzelnen Federästen zusammengesetzt.
Mit intersexueller Selektion erklärbar?
Die Schwanzfedern mit ihrem ausgefeilten Augenmuster spielen zweifellos eine Rolle im Balzritual. Ob Weibchen aber von ihnen „angezogen“ werden oder sie nur nüchtern als Signal zu Paarungsbereitschaft des Männchens interpretieren, ist eine andere Frage.
Selbstverständlich geht die Schönheit der Schwanzfedern weit über das hinaus, was für ein Signal zur Paarungsbereitschaft erforderlich wäre. Doch daraus folgt nicht objektiv zwingend eine anziehende Wirkung bei den Weibchen, denn man könnte ebenso argumentieren, dass die Balz als ein "schöner" Prozess ein geeigneter Ort für überflüssige geschaffene Schönheit ist.
Interessanterweise gibt es zu der Frage, ob die Pfauenhenne tatsächlich von den prächtigen Schwanzfedern angezogen wird, gegenteilige Studien, wie Woetzel (2017) bemerkt. Während beispielsweise Takahashi et al. (2008) in einer Studie keine Präferenz der Pfauenweibchen für längere Schwanzfedern oder eine höhere Anzahl „Augen“ nachweisen konnten, beobachteten Yorzinski et al. (2013), dass Pfauenhennen ein besonderes Augenmerk auf den unteren Teil des Federschwanzes legen.
Die wohl größte Herausforderung für eine evolutionäre Erklärung der Schwanzfedern des männlichen Pfaus liegt aber in den feinen und subtilen Details. Während Pfauen durchaus noch in der Lage sein mögen, offensichtlichere Details wie die Anzahl der Augen-Federn zu erkennen, gibt es keinen Nachweis, dass Pfauenhennen auch subtile Details wie die braune Färbung des Federschaftes am Ende registrieren können (Burgess 2023; Woetzel 2017).
Da Pfauenhennen diese feinen Details also wohl auch nicht bevorzugen können, hätten sie evolutiv ohne Selektionsdruck auch nicht entstehen können – jedenfalls, solange sie nicht genetisch mit anderen Merkmalen gekoppelt sind (Pleiotropie = der Effekt, wenn ein Gen mehrere Merkmale steuert). Burgess (2023) benennt und diskutiert noch weitere Probleme.
Wenn man die hochgradige Ordnung und Präzision sowohl des Gesamtbildes (geometrische Anordnung der Federn) als auch der Details (z.B. Dünnschichtinterferenz) betrachtet, liegt es auf der Hand, dass solche feinen, abgestimmten Strukturen eher bei einem Designer zu erwarten sind als bei ungerichteten und ziellosen Naturprozessen.
Das Prachtgefieder des männlichen Pfau kann durch sexuelle Selektion nicht plausibel erklärt werden, es ist und bleibt ein Paradebeispiel des Schöpfungsindizes der unnötigen Schönheit.
Und was ist mit intrasexueller Selektion?
Dem aufmerksamen Leser wird aufgefallen sein, dass sich die Kritik in diesem Artikel vornehmlich auf die intersexuelle Selektion bezieht. Dies liegt daran, dass sie eben jene extravaganten Merkmale erklären soll, die von schöpfungswissenschaftlicher Seite als Schöpfungsindizien der unnötigen Schönheit betrachtet werden.
Sekundäre Geschlechtsmerkmale der intrasexuellen Selektion spielen deutlich seltener eine Rolle in Argumenten für Schöpfung. Die Erhaltung und Ausbreitung intrasexueller Merkmale, die einen Vorteil im Wettbewerb um den Zugang zu Paarungspartnern ermöglichen, sind insofern nachvollziehbar, als hier ein plausibler Selektionsdruck von Beginn an angebracht werden kann. Dies erklärt zwar nicht den Ursprung komplexer Strukturen, wohl aber deren Erhalt oder Vergrößerung.
Doch auch wenn intrasexuelle Selektion evolutionstheoretisch plausibler ist, spricht sie nicht gegen einen Schöpfer. Denn die schöpfungstheoretische Deutung dieser Merkmale als für den Wettkampf geschaffen ist nicht weniger plausibel.
Herausforderung und Deutung aus Schöpfungssicht:
Auch wenn Vertreter der Evolutionstheorie mit einigen schwerwiegenden Problemen zu kämpfen haben, stellt sich eine Frage auch unter Zugrundelegung des biblischen Schöpfungsberichtes: Wenn die belebte Schöpfung seit dem Sündenfall degeneriert, warum existiert dann überhaupt noch ein derart großes Ausmaß an unnötiger, aber in der gefallenen Schöpfung nachteiligen Schönheit?
Wie bereits erwähnt, kann man die weibliche Bevorzugung als geschaffenen Erhaltungsmechanismus deuten, wie es Burgess (2023) tut. Da Gene, die eine Präferenz für Merkmale der Fortpflanzungspartner (= Präferenzgene) codieren, auch degenerieren können, geht Woetzel (2017) noch einen Schritt weiter und stellt die Hypothese auf, dass die Gene für extravagante Merkmale mit Genen für überlebenswichtige Funktionen oder Merkmale verbunden sein könnten. Wenn dann die Gene für ein intersexuelles Merkmal mutationsbedingt beschädigt werden, hätte das auch negative Auswirkungen auf lebensnotwendige Strukturen. So würde der Verlust unnötiger Schönheit durch natürliche Selektion zusätzlich verlangsamt.
Woetzel (2017) nennt es zwar nicht explizit, doch dieses Prinzip wäre natürlich auch auf Präferenzgene anwendbar. In dem Fall würde eine Verbindung mit weiteren Genen Präferenzgene vor Degeneration schützen und damit indirekt auch das entsprechende Schmuckmerkmal.
Woetzel (2017) schlägt nun vor, die Hypothese für genetisch gekoppelte Schmuckmerkmale mit wichtigen Lebensfunktionen mittels Knockout-Experimenten zu testen: Wenn Gene für ein für die Fitness nachteiliges Schmuckmerkmal identifiziert werden, könnte man diese ausschalten und beobachten, ob negative Auswirkungen auf weitere Funktionen / Merkmale auftreten.
Tatsächlich wurden die Vorhersagen dieser Hypothese mehrfach in Studienergebnissen bestätigt. Zwei Beispiele dazu:
1) Männchen von Drosophila melanogaster (Fruchtfliege) besitzen an den Vorderbeinen kleine, bürstenartige Strukturen, sogenannte Geschlechtskämme. Mit diesen halten sie sich während der Paarung am Hinterleib des Weibchens fest. Diese Geschlechtskämme sind schwarz gefärbt, was an sich keinen funktionellen Nutzen hat, im Gegenteil, es erhöht die Sichtbarkeit für Fressfeinde. Somit kann die schwarze Färbung als Schmuckmerkmal verstanden werden.
Massey et al. (2019) identifizierten das yellow-Gen als verantwortlich für die schwarze Färbung der Geschlechtskämme, weil dieses für die Produktion von schwarzem Melanin codiert. Massey et al. (2019) schalteten das Gen bei genetisch veränderten Fruchtfliegen aus. Die Folge war nicht nur eine gelbe Farbe der Geschlechtskämme, sondern auch, dass diese weicher wurden. In der Folge hatten die betreffenden Männchen nur noch einen Paarungserfolg von 3%. Das Gen und somit das Melanin für die an sich funktionslose schwarze Farbe ist also auch für die erforderliche Härte einer für den Geschlechtsakt benötigten Struktur relevant.
2) Rossi et al. (2024) identifizierten in drei Arten von Heliconius-Schmetterlingen eine Genomregion, die mit der männlichen Bevorzugung roter beziehungsweise weißer Flügelmuster bei Weibchen in Verbindung gebracht wird. Eines dieser Gene wird je nach Art und entsprechend unterschiedlicher Präferenz unterschiedlich exprimiert (= ausgeprägt). Rossi et al. (2024) entfernten einen Teil des proteincodierenden Bereiches dieses Gens in einer der Arten. In der Folge wiesen die betreffenden männlichen Schmetterlinge nicht nur ein verändertes Balzverhalten auf, teilweise kam es sowohl vor als auch nach dem Schlüpfen zu einer verminderten Lebensfähigkeit. Zusätzlich waren einige der veränderten Männchen nicht mehr in der Lage, zu fliegen.
Woetzels Kopplungs-Hypothese wurde somit in diesen Fällen bestätigt. Die Erhaltung unnötiger, z. T. nachteilhafter Schönheit in einer von Degeneration bedrohten Schöpfung kann durch sexuelle Selektion und durch eine Verbindung der Gene für entsprechend extravagante Merkmale, bzw. deren Bevorzugung mit anderen, wichtigeren Merkmalen bzw. Funktionen erklärt werden.
Fazit:
Sowohl inter- als auch intrasexuelle Selektion als gegenwärtiger, erhaltender Prozess sind empirisch gesichert. Es ist häufig beobachtet worden, dass bestimmte sekundäre Geschlechtsmerkmale den Fortpflanzungserfolg steigern und diese dadurch in der Population erhalten werden.
Die erstmalige Entstehung dieser Merkmale – insbesondere der unnötig schönen bzw. aufwändigen – durch sexuelle Selektion ist dagegen erheblich problematischer. Abgesehen von Einzelfällen, in denen die Sensory-exploitation-Hypothese (d. h. sexuelle Präferenzen basieren auf anderen Präferenzen) greifen könnte, gibt es keine plausible naturalistische Erklärung, ob zuerst das neue Merkmal oder seine Bevorzugung evolvierte und wie das eine in der Zwischenzeit ohne das andere erhalten bleiben konnte. Auch Erklärungen für den Grund der weiblichen Bevorzugung extravaganter, nachteiliger Merkmale sind nicht plausibler.
Die Theorie der sexuellen Selektion wurde von Charles Darwin eingeführt, weil er eben jene unnötig schönen und aufwändigen Merkmale, die auf ein Schöpfer deuten, nicht mit natürlicher Selektion erklären konnte. Doch dieser Ansatz ist bislang gescheitert, denn auch diese Form der Selektion kann einen Schöpfer nicht ersetzen.
Zugleich sind die beobachtbaren Aspekte der sexuellen Selektion problemlos im Schöpfungsmodell als geschaffener Mechanismus zur Erhaltung geschaffener Extravaganz deutbar (Burgess 2023; Woetzel 2017).
Danksagung:
Ich möchte mich herzlich bei Dr. Reinhard Junker, Dr. Boris Schmidtgall, Benjamin Scholl und Dr. Peter Borger für das Korrekturlesen dieses Artikels bedanken. Es gab viele hilfreiche Hinweise und manche gute Diskussion. Ebenfalls danken möchte ich Prof. Stuart Burgess für seine Erlaubnis einige seiner Bilder zu verwenden.
Literatur:
Bergman J. (1996) The Enigma of Sex and Evolution. Creation Research Society Quarterly, 33 (3): 7.
Bergman J. (2004) Problems in sexual selection theory and neo-Darwinism. TJ, 18(1), S. 112–119.
Bergman J. (2005) Darwinism and the Deterioration of the Genome. Creation Research Society Quarterly 42(2), S. 104–114.
Bolten A. (2020) Werbeverhalten mit Lebensgefahr. scinexx. https://www.scinexx.de/dossierartikel/werbeverhalten-mit-lebensgefahr/
Brennan P. L. R. et al. (2007) Coevolution of Male and Female Genital Morphology in Waterfowl. PLoS ONE.
Burgess S. (2023) The beauty of the peacock tail and the problems with the theory of sexual selection. creation.com. Erstveröffentlichung 2001 im Journal of Creation 15(2).
Cassini M. (2020) Sexual aggression in mammals. Mammal Review. 51. 10.1111/mam.12228.
Clutton-Brock T. H. & Parker G. A. (1992) Potential reproduction rates and the operation of sexual selection. The Quarterly Review of Biology 67(4):437-456.
Darwin C. (1871) The descent of man and selection in relation to sex. London: Murray.
Fisher R. A. (1930) The genetical theory of natural selection. Clarendon Press.
Grose J. (2011) Modelling and the Fall and Rise of the Handicap Principle. Biology & Philosophy 26 (5): 677–696.
Head M. L. & Brooks R. (2006) Sexual coercion and the opportunity for sexual
selection in guppies. ANIMAL BEHAVIOUR, 2006, 71, 515–522, doi:10.1016/j.anbehav.2005.04.017.
Kane P. & Zollman K. J. S. (2015) An Evolutionary Comparison of the Handicap Principle and Hybrid Equilibrium Theories of Signaling. PloS ONE 10 (9): e0137271.
Kirkpatrick M. (1986) The Handicap Mechanism of Sexual Selection Does Not Work. The American Naturalist 127 (2): 222–240.
Lee, P. L. M. & Hays G. C. (2004). Polyandry in a marine turtle: females make the best of a bad job. PNAS.
Madden J. R. & Tanner K. (2003) Preferences for coloured bower decorations can be explained in a non-sexual context. Animal Behaviour, 65(6), 1077–1083. https://doi.org/10.1006/anbe.2003.2126
Massey J. H. et al. (2019) The yellow gene influences Drosophila male mating success through sex comb melanization eLife 8:e49388.
Natterer P. (2010) Motoo Kimura und John Sanford zur Neutralen Theorie der molekularen Evolution und zur These der genetischen Entropie. Wissenschaftspublizistik novum studium generale — Liberal Education Publishing & Consulting.
Ritchie M. G. (2007) Sexual selection and speciation. Annu Rev Ecol Evol Syst. 38:79–102.
Rodd F. H. et al. (2002): A possible non-sexual origin of mate preference: are female guppies attracted to orange-colored objects because they resemble fruits? Proceedings of the Royal Society of London. Series B: Biological Sciences, 269(1490), 475-481.
Rosenthal G. G. (2017) Mate Choice: the evolution of sexual decision making from microbes to humans. Princeton: Princeton University Press.
Rossi M. et al. (2024) Adaptive introgression of a visual preference gene. Science 383,1368-1373. DOI:10.1126/science.adj9201
Roughgarden J. & Akçay E. (2010) Do We Need a Sexual Selection 2.0?” Animal Behaviour 79 (3): e1–e4.
Ryan M. J. (1990) Sexual selection, sensory systems and sensory exploitation. Oxford Survey of Evolutionary Biology 7, S. 156-195.
Schloegl C. & Fischer J. (2017) Causal Reasoning in Non-Human Animals, in Michael R. Waldmann (ed.), The Oxford Handbook of Causal Reasoning, Oxford Library of Psychology (online edn, Oxford Academic, 10 May 2017), https://doi.org/10.1093/oxfordhb/9780199399550.013.36.
Shuker D. M. & Kvarnemo C. (2021) The definition of sexual selection, Behavioral Ecology. Volume 32, Issue 5, S. 781–794, https://doi.org/10.1093/beheco/arab055
Simmons L.W. & Wedell N. (2020) Fifty years of sperm competition: the structure of a scientific revolution. Philos Trans R Soc Lond B Biol Sci 7; 375 (1813): 20200060. https://doi.org/10.1098/rstb.2020.0060
Scholes E. & Laman T. G. (2018) Distinctive courtship phenotype of the Vogelkop
Superb Bird-of-Paradise Lophorina niedda Mayr, 1930 confirms new species status.
PeerJ 6:e4621; DOI 10.7717/peerj.4621.
Smuts B. B. & Smuts R. W. (1993) Male aggression and sexual coercion of females in nonhuman primates and other mammals: evidence and theoretical implications. Advances in the Study of Behavior 22: 1–63.
Számadó S. et al. (2026) A general signalling theory: why honest signals are explained by trade-offs rather than costs or handicaps. Journal of Evolutionary Biology 39, 171–189.
Takahashi M. et al. (2008) Peahens do not Prefer Peacocks With More Elaborate Trains. Animal Behaviour 75 (4): 1209–1219.
Wigby S. & Chapman T. (2004) Sperm competition, Current Biology. Volume 14, Issue 3, R100 - R103.
Witte K. (2009) Sexuelle Selektion – Die Bedeutung genetischer und sozialer Faktoren für die weibliche Partnerwahl. PdN-BioS, 58(3), S. 18–22.
Woetzel D. (2017) The Origin of Beauty: Contrasting the Handicap Hypothesis and Design Theory. Answers Research Journal, 10, 285–292.
https://answersresearchjournal.org/origin-beauty-handicap-hypothesis-design/.
Yorzinski J. L. et al. (2013) Through Their Eyes: Selective Attention in Peahens During Courtship. Journal of Experimental Biology 216 (16): 3035–3046.
Zahavi A. (1975) Mate selection - a selection for a handicap. Journal of theoretical Biology 53, S. 205-214.
[Eine ähnliche Version dieses Artikels ist auch in drei Teilen bei genesis-net erschienen.]



Kommentare